How can we accurately classify people in a manner that corresponds to real biological differences rather than culturally defined stereotypes. The answer to this question is not simple. There are three basic ways in which anthropologists have tried to do it in the past. These are generally referred to as the typological, populational, and clinal models.
Typological
Model
During the 19th and early 20th centuries,
anthropologists and biologists naively divided all people into
distinct geographical groups on the basis of what
they saw as the regular occurrence together of selected traits. They were using the
typological model
![]() which has the same basis as the
commonly used categories of race today. This approach focuses on a
small number of traits that are readily observable from a distance such as skin color,
hair form, body build, and stature. Inherent in the
typological model is the notion that there have been "pure" unmixed races in
the past. The roots of
this model for classifying people go back at least to the 18th
century Swedish naturalist, Carolus Linnaeus. He proposed the existence
of four biological subspecies or races of humans corresponding to geographic
regions:
which has the same basis as the
commonly used categories of race today. This approach focuses on a
small number of traits that are readily observable from a distance such as skin color,
hair form, body build, and stature. Inherent in the
typological model is the notion that there have been "pure" unmixed races in
the past. The roots of
this model for classifying people go back at least to the 18th
century Swedish naturalist, Carolus Linnaeus. He proposed the existence
of four biological subspecies or races of humans corresponding to geographic
regions:
| Homo sapiens Eoropeus albescens | ("white" people from Europe) |
| Homo sapiens Africanus negreus | ("black" people from Africa) |
| Homo sapiens Asiaticus fucus | ("dark" people from Asia) |
| Homo sapiens Americanus rubescens | ("red" people from the Americas) |
The typological model is based on what is now known to be a false assumption concerning the nature of human variation--that is that we can be unambiguously assigned to a "race" on the basis of selected anatomical traits. In fact, when we look at specific individuals, we often run into difficulty trying to categorize them. For example, on the basis of skin color, we might put them into one "race" and on the basis of nose shape, body form, or blood type, they might go into others.
Even if we base the classification on a single characteristic, the typological model fails because most biological traits are continuous variables. That is to say, there is a progressive range of variation that our preconceptions sometimes prevent us from seeing. The typological model causes us to lump people together into arbitrarily assigned categories. For instance, we tend to classify people as being slender, medium build, or fat despite the fact that there is a continuum of body weight and size among humans. The same is true of skin color and other commonly used "racial" characteristics.
The typological model usually leads us to wrongly assume that the people within any "race" are genetically and anatomically more alike than they are like people from other "races." This all depends on the traits that are being compared. For instance, many Americans believe that people of African descent have broad noses. In fact, both the widest and the narrowest noses are found among the people of that continent. Recent research comparing human DNA sequences from around the world has shown that 90% of human genetic variation exists within what we have previously assumed to be more or less separate "races" and only 10% between them. In other words, "racial" groups are far from being homogenous.
Another major problem with the typological
model is that the number of "races" you end up with depends on the number and
kinds of traits employed in the classification. The more traits used, the fewer
people in the world there are who share them. For example, light skin color is
considered to be a defining characteristic of Europeans. However, when you add the
criteria of narrow noses, straight hair, and tall stature, many Europeans would be
excluded altogether or the European racial category would have to be further subdivided
into several smaller "races." Since the number of "races" can be so
easily changed by the way they are defined, it is clear that they do not really
exist as distinct biological groupings of people. Instead, they are arbitrary
creations that reflect our ethnocentric
![]() views of ourselves and other
people. They are mainly cultural rather than
biological groupings.
views of ourselves and other
people. They are mainly cultural rather than
biological groupings.
Populational
Model
By the early
1940's, most biological anthropologists recognized
these problems and adopted the populational model
![]() as an alternative. This
emerged from perspectives developed by
population genetics. It
is based on the idea that the only significant groups, in evolutionary terms,
consist of people whose ancestors have more or less exclusively mated with
each other for thousands of years. Individuals in such distinct breeding populations would be expected
to share many genetically inherited traits and to have a similar appearance.
as an alternative. This
emerged from perspectives developed by
population genetics. It
is based on the idea that the only significant groups, in evolutionary terms,
consist of people whose ancestors have more or less exclusively mated with
each other for thousands of years. Individuals in such distinct breeding populations would be expected
to share many genetically inherited traits and to have a similar appearance.
This approach to understanding the patterns of human biological diversity differs radically from the typological model. The latter starts by defining traits that presumably characterize a "race" and then looks around the world to see who has them. In contrast, the populational model looks for breeding populations first and then considers the anatomical and physiological traits that may distinguish them.
While the populational approach makes theoretical sense, it is undermined by the fact that throughout history humans have rarely mated within a single group for long. Cultural and geographic barriers to intergroup mating have most often broken down over time. Speeding up this process of intermixture of humanity have been the mass intercontinental migrations of the last 500 years. This process is actually accelerating today largely due to relatively inexpensive and easy intercontinental travel. Biological anthropologists have found only a few moderately distinct breeding populations still existing. As a result, the populational model is of little help in understanding most of human variation today. However, it is of value in studying the few relatively isolated communities that have survived.
Clinal
Model
By the
early 1960's, sufficient data
had been gathered for biological anthropologists to understand that a
clinal model
![]() more accurately reflects the
true nature of human biological variation. This model is based on the fact that
genetically inherited traits most often change
gradually in frequency from one geographic
area to another. For instance, the allele for type B blood generally increases from west to east in
Europe. We can record different frequency zones, or clines
more accurately reflects the
true nature of human biological variation. This model is based on the fact that
genetically inherited traits most often change
gradually in frequency from one geographic
area to another. For instance, the allele for type B blood generally increases from west to east in
Europe. We can record different frequency zones, or clines
![]() (as shown in the map below). Unlike the
typological and populational models, the clinal model does not result in the definition of
distinct groups or races of people.
(as shown in the map below). Unlike the
typological and populational models, the clinal model does not result in the definition of
distinct groups or races of people.
|
Clinal distribution of
the |
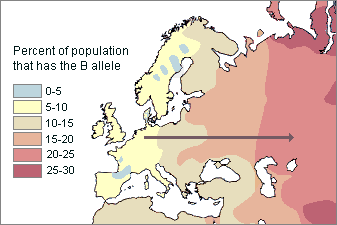 |
Gradual changes in gene frequency from one region to another are mostly due to the simple fact that the chance of our mating with someone is usually directly related to the distance they live from us. People whose ancestors have lived close to ours for many generations are more likely to share genetically inherited traits with us than are people who live further away. However, as long distance transportation systems have become more accessible and dependable, the distance from home that we travel and potentially find mates has increased. Despite this change, most people still usually end up marrying others who live within a few hundred miles of their home.
Unfortunately, the pattern of human
variation around the world cannot be entirely understood by the clinal model alone.
The distribution of some traits is partly discontinuous
![]() . The example of red hair in England
(described in the first section of this tutorial) is not unique. There
are other traits that have non-clinal distribution patterns. These also can
be understood as results of historical migrations or exclusive breeding within more or less closed
communities. For example, the map below shows that the frequency of people
who have the B blood allele generally increases from east Asia to
central Asia. Within this more or less continuous cline, there are isolated pockets of relatively low B
allele frequency. Therefore, the distribution of this genetically inherited trait appears
to be mostly clinal, but it is also partly
discontinuous.
. The example of red hair in England
(described in the first section of this tutorial) is not unique. There
are other traits that have non-clinal distribution patterns. These also can
be understood as results of historical migrations or exclusive breeding within more or less closed
communities. For example, the map below shows that the frequency of people
who have the B blood allele generally increases from east Asia to
central Asia. Within this more or less continuous cline, there are isolated pockets of relatively low B
allele frequency. Therefore, the distribution of this genetically inherited trait appears
to be mostly clinal, but it is also partly
discontinuous.
|
Clinal and discontinuous |
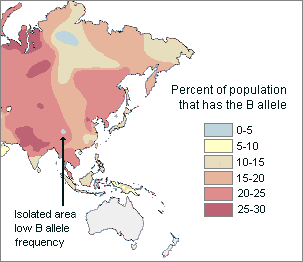 |
How
Can Human
Variation Be Best Described?
It is clear that all of the models fail to adequately carry out the entire job. The typological model is the most unsound because presumed racial traits are not found exclusively within defined races. In addition, focusing on new sets of traits often results in assigning people to different races, despite the fact that they were lumped into the same race before. The populational model makes sense theoretically but fails to account for most of the distribution patterns around the world because we do not limit our breeding to isolated populations. The clinal model comes the closest to grasping the real nature of human variation. However, it is undermined by the occasional discontinuous distribution resulting from migrations and the few remaining small isolated communities. The patterns of human variation around the world are not only highly complex but also are constantly shifting through time. Furthermore, the rate of change in the patterns has been accelerating as our numbers grow and as long distance travel and migration become more routine. Contributing to the intermixture of peoples in the Western World has been a reduction of inhibitions about marriage across perceived "racial" lines.
In the final analysis, it is important to keep in mind that all humans around the world today are biologically quite similar despite our superficial differences. In fact, we apparently are 99.9% genetically identical. Most of the differences between us are due to our unique individual traits and being male or female. When we are compared to many other kinds of animals, it is remarkable how little variation exists within our own species. There is 2-3 times more genetic variation among chimpanzees, 8-10 times more among orangutans, and thousands of times more in many insect species. Most biological anthropologists would agree that human variation is not now sufficient to warrant defining separate biological races, varieties, or sub-species. However, it very likely was in our distant prehistoric past.
In order to better understand the true patterns of human variation, biological anthropologists have gathered detailed data about genetically inherited traits. Most of this work has been done with blood typing, but DNA sequence comparisons are now providing an even more detailed understanding of our human biological diversity.
Genetic Distances Between Human Populations Today
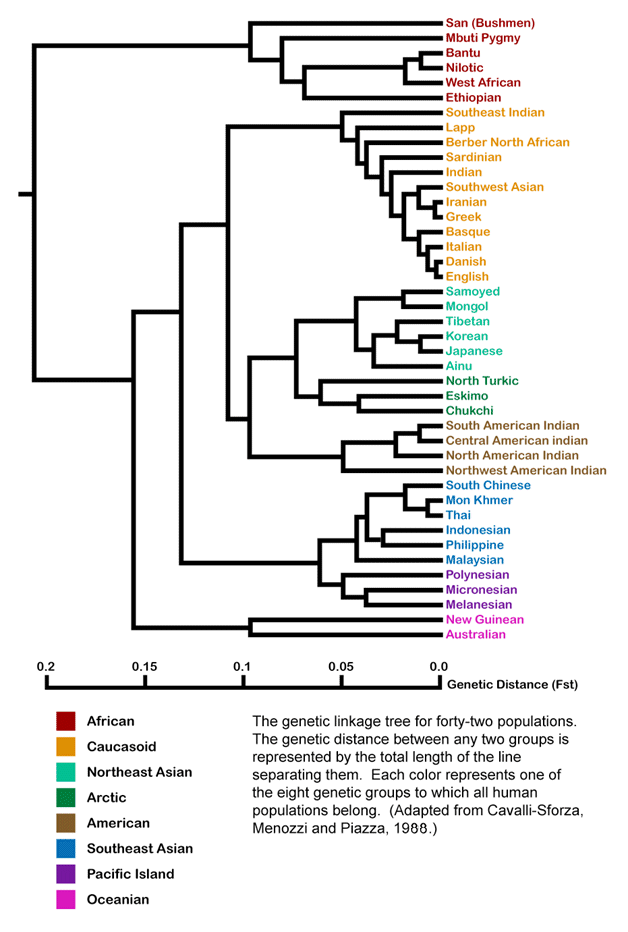
These DNA sequence comparisons indicate that the farther apart populations are from each other geographically, the less similar they are genetically. This is likely due to the fact that people from more distant populations are less likely to have the opportunity to mate together than they are with their close neighbors.